Regulering van cholesterolsintese
Die regulering van die sintese van cholesterol - die sleutelensiem daarvan (HMG-CoA-reduktase) word op verskillende maniere uitgevoer.
Fosforylering / defosforylering van HMG-reduktase. Met die toename in die insulien / glukagon-verhouding fosforylateer hierdie ensiem en beweeg dit in 'n aktiewe toestand. Die werking van insulien word deur 2 ensieme uitgevoer.
HMG-CoA reduktase kinase fosfatase, wat die kinase omskep in 'n onaktiewe gedefosforileerde toestand:
Fosfotase HMG-CoA reduktase deur dit in 'n gedefosforyleerde aktiewe toestand om te skakel. Die resultaat van hierdie reaksies is die vorming van 'n gedefosforyleerde aktiewe vorm van HMG-CoA-reduktase.
Gevolglik neem cholesterol toe gedurende die absorpsietydperk. Gedurende hierdie periode neem die beskikbaarheid van die aanvanklike substraat vir die sintese van cholesterol - asetiel-CoA toe (as gevolg van die eet van voedsel wat koolhidrate en vette bevat, aangesien CoA-asetiel gevorm word tydens die afbreek van glukose en vetsure).
In die postabsorberende toestand stimuleer glukagon deur proteingenase A die fosforylering van HMG - CoA - reduktase, wat dit in 'n onaktiewe toestand maak. Hierdie werking word versterk deurdat glukagon terselfdertyd die fosforilering en die inaktivering van die fosfotase van HMG-CoA-reduktase stimuleer, en sodoende die HMG-CoA-reduktase in 'n gefosforyleerde onaktiewe toestand gehou word. As gevolg hiervan, word die sintese van cholesterol in die postabsorpsieperiode en tydens vas geïnhibeer deur endogene sintese. As die cholesterolinhoud in voedsel op 2% gebring word, neem die sintese van endogene cholesterol skerp af. Maar die volledige staking van cholesterol-sintese kom nie voor nie.
Die mate van inhibisie van cholesterolbiosintese onder invloed van cholesterol wat van voedsel afkomstig is, wissel van persoon tot persoon. Dit dui op die individualiteit van die prosesse van cholesterolvorming. Deur die intensiteit van cholesterolsintese te verminder, is dit moontlik om die konsentrasie in die bloed te verminder.
As die balans tussen die inname van cholesterol met voedsel en die sintese daarvan in die liggaam aan die een kant en die uitskeiding van galsure en cholesterol aan die ander kant verbreek word, verander die konsentrasie van cholesterol in die weefsel en bloed. Die ernstigste gevolge hou verband met 'n toename in bloedkolesterolkonsentrasie (hipercholesterolemie), terwyl die waarskynlikheid om aterosklerose en cholelithiasis te ontwikkel, verhoog.
Gesinshipercholesterolemie (HCS) - hierdie vorm kom baie meer voor - ongeveer 1 pasiënt per 200 mense. 'N Geërfde defek in HCS is 'n skending van die opname van LDL deur selle, en dus 'n afname in die tempo van LDL-katabolisme. As gevolg hiervan styg die konsentrasie van LDL in die bloed, sowel as cholesterol, aangesien daar baie daarvan in LDL is. Daarom is die afsetting van cholesterol in weefsels, veral in die vel (xanthomas), in die wande van arteries kenmerkend met HCS.
Inhibering van die sintese van HMG-CoA-reduktase
Die finale produk van die metaboliese metaboliese weg. Dit verminder die tempo van transkripsie van die HMG-CoA-reduktase-geen, en belemmer dus sy eie sintese. Die lewer sintetiseer galsure aktief van cholesterol, en die remming van die galsure is dus die aktiwiteit van die HMG-CoA-reduktase-geen. Aangesien HMG-CoA-reduktase na sintese van ongeveer 3 bestaan, is die inhibering van die sintese van hierdie cholesterolensiem 'n effektiewe regulering.
Het u nie gevind waarna u gesoek het nie? Gebruik die soektog:
Cholesterol ester ruil
Die cholesterolfonds bevat gratis cholesterol- en cholesterolesters, wat in selle en in bloedlipoproteïene aangetref word.
Deel II Metabolisme en energie
In selle vind die verestering van cholesterol plaas met die werking van acyl-CoA-cholesterol-acyltransferase (AChAT):
Acyl-CoA + Cholesterol - * HS-KoA + Acylcholesterol
In menslike selle word linoleylcholesterol hoofsaaklik gevorm. Anders as gratis cholesterol, is die esters daarvan in selmembrane in baie klein hoeveelhede vervat en word dit hoofsaaklik in die sitosol as deel van lipiedruppels aangetref. Die vorming van esters kan enersyds beskou word as 'n meganisme om oortollige cholesterol uit die membrane te verwyder, en andersyds as 'n meganisme vir die opberg van cholesterol in die sel. Hulpbronmobilisering vind plaas met die deelname van esterase-ensieme wat cholesterolesters hidroliseer:
Asielcholesterol + H 2O - * Vetsuur + cholesterol
Die sintese en hidrolise van esters kom in baie selle voor, maar is veral aktief in selle van die bynierskors: in hierdie selle word tot 80% van alle cholesterol deur esters voorgestel, terwyl dit in ander selle gewoonlik minder as 20% is.
Die vorming van esters in bloedliponroteïne vind plaas met die deelname van lesitien-cholesterol-asieltransferase (LHAT), wat die oordrag van die asielresidu van die i-posisie van lesitien na cholesterol kataliseer (Fig. 10.31). LHAT word in die lewer gevorm, in die bloed afgeskei en aan lipoproteïne geheg. Die esterifikasietempo vir verskillende lipoproteïene is beduidend verskillend en hang af van die teenwoordigheid van apolipoproteïene wat LHAT (hoofsaaklik apo-AT, sowel as CI) aktiveer of hierdie ensiem inhibeer (C-II). Die aktiefste LHAT in HDL, waarin apo-AT moet olee 2/3 van alle proteïene. Die grootste coli honneurs gevorm esters van olie en linoleïensuur sure. Die ander lipoproteïene ester vorming plaasvind teen 'n stadiger koers as in HDL.
Fig. 10,31. Die vorming van cholesterolesters onder die werking van LHAT
LHAT is gelokaliseer in die oppervlaklaag van HDL en gebruik cholesterol in die fosfolipied monolag as substraat. Die cholesterolesters wat hier gevorm word, word as gevolg van hul volledige hidrofobisiteit swak in die grond gehou
Hoofstuk 10. Metabolisme en lipiedfunksie
fosfolipied monolag en gedompel in die lipiedkern van die lipoproteïen. Terselfdertyd word 'n plek vir cholesterol in die fosfolipied monolag vrygestel, wat gevul kan word met cholesterol uit selmembrane of uit ander lipoproteïene. HDL blyk dus 'n cholesterolval te wees as gevolg van die werking van LHAT.
Galsuur sintese
In die lewer word 'n deel van die cholesterol omgesit in galsure. Galliensure kan beskou word as afgeleides van cholansuur (Fig. 10.32).
Cholansuur as sodanig word nie in die liggaam gevorm nie. By hepatosiete produseer cholesterol direk chenodeoksigoliese en cholesure - primêre galsure (Fig. 10.33, sien ook Fig. 10.12).
Cholesterolbiosintese
Cholesterolbiosintese kom voor in die endoplasmiese retikulum. Die bron van alle koolstofatome in die molekule is asetiel-SCoA, wat hier kom uit mitochondria as deel van sitraat, soos in die sintese van vetsure. Die cholesterolbiosintese verbruik 18 ATP-molekules en 13 NADPH-molekules.
Die vorming van cholesterol kom voor in meer as 30 reaksies, wat in verskillende fases gegroepeer kan word.
1. Sintese van mevalonzuur.
Die eerste twee sintesereaksies val saam met die ketogenesereaksies, maar na die sintese van 3-hidroksy-3-metielglutaryl-ScoA, gaan die ensiem binne hidroksimetiel-glutaryl-ScoA-reduktase (HMG-SCOA-reduktase), wat mevalonzuur vorm.
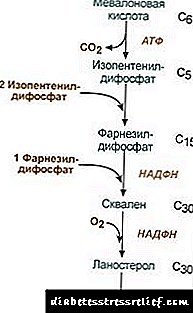 |
Cholesterol sintese reaksieskema2. Sintese van isopentenyldifosfaat. Op hierdie stadium word drie fosfaatreste aan mevalonzuur vasgemaak, dan word dit gekarboksileer en ontwater. 3. Nadat die drie molekules isopentenyldifosfaat gekombineer is, word farnesyldifosfaat gesintetiseer. 4. Die sintese van squaleen vind plaas wanneer twee farnesyldifosfaatreste gebind word. 5. Na ingewikkelde reaksies fietseer lineêre squaleen na lanosterol. 6. Die verwydering van oortollige metielgroepe, herstel en isomerisering van die molekule lei tot die voorkoms van cholesterol. Regulering van die aktiwiteit van hidroksimetylglutaryl-S-CoA-reduktase3. Die tempo van cholesterolbiosintese hang ook af van die konsentrasie spesifieke draproteïenvoorsiening te maak vir die binding en vervoer van hidrofobiese intermediêre sintese-metaboliete. U kan u mening vra of laat. Die belangrikste punt van regulering is die reaksie van die vorming van mevalonzuur. 1. Allosteriese regulering. Cholesterol, en in die lewer - en galsure rem die HMG-CoA-reduktase. 2. Onderdrukking van die sintese van HMG-CoA reduktase cholesterol. 3. Regulering deur fosforylering-de-fosforylering van HMG-CoA-reduktase, aktiewe nie-fosforileerde vorm. Glukagon veroorsaak deaktivering, en insulien veroorsaak aktivering deur 'n ingewikkelde reaksie. Dus verander die tempo van cholesterol sintese met 'n verandering in absorberende en postabsorpsietoestande. 4. Die tempo van sintese van HMG-CoA-reduktase in die lewer is onderhewig aan duidelike daaglikse skommelinge: maksimum om middernag en minimum in die oggend. Cholesterol ester ruil In die selle verestering van cholesterol vind plaas wanneer dit blootgestel word asiel-KoA cholesterol acyltransferase (ACAT): Acyl-CoA + Cholesterol ® NS-CoA + Acylcholesterol In selle word hoofsaaklik linoleylcholesterol gevorm. Esters kom hoofsaaklik in die sitosol voor as deel van lipieddruppels. Die vorming van esters kan enersyds beskou word as 'n meganisme om oortollige cholesterol uit die membrane te verwyder, en andersyds as 'n meganisme om cholesterol in die sel te stoor. Die mobilisering van reserwes vind plaas met die deelname van ensieme esterasehidroliseer cholesterolesters: Asielcholesterol + H2O ® vetsuur + cholesterol Die sintese en hidrolise van esters is veral aktief in die selle van die bynierskors. In bloedlipoproteïene estervorming vind plaas met die deelname van lesitien-cholesterol acyltransferase (LHAT), wat die oordrag van die asielresidu van lesitien na cholesterol kataliseer. LHAT word in die lewer gevorm, in die bloedstroom afgeskei en aan lipoproteïene geheg. Die aktiefste LHAT in HDL, waar dit gelokaliseer word in die oppervlaklaag. Die cholesterolesters wat hier gevorm word, is hidrofobies en gedompel in die lipiedkern. In die fosfolipiede monolaag is daar vrye ruimte vir cholesterol, wat gevul kan word met cholesterol uit selmembrane of van ander lipoproteïene. HDL blyk dus 'n cholesterolval te wees as gevolg van die werking van LHAT. Galsuur sintese In die lewer word 'n deel van die cholesterol omgesit in galsure. Galsure kan beskou word as afgeleides van cholansuur. Cholansuur as sodanig word nie in die liggaam gevorm nie. In hepatosiete van cholesterol word primêre galsure gevorm - chenodeoxycholic en cholic. Na die afskeiding van gal in die ingewande onder die werking van ensieme van die dermflora, word sekondêre galsure daaruit gevorm - lithocholic en deoxycholic. Dit word in die ingewande opgeneem, met die bloed van die portale aar in die lewer en dan in die gal. Gal bevat hoofsaaklik gekonjugeerde galsure, dit wil sê hul verbindings met glisien of taurien. Die konsentrasie van galsure in gal is ongeveer 1%. Die grootste deel van galsure is betrokke by hepatoenteriese sirkulasie.'N Klein gedeelte van galsure - ongeveer 0,5 g per dag - word in ontlasting uitgeskei. Dit word vergoed deur die sintese van nuwe galsure in die lewer; die galsuurfonds word binne tien dae opgedateer. Cholesterol word ook hoofsaaklik deur die ingewande geskei. Dit kom in die ingewande in met voedsel en van die lewer as deel van gal. Cholesterol wat in die bloed opgeneem is, bevat 'n fraksie wat uit gal voorkom (endogene cholesterolgesintetiseer in die lewer), en die fraksie afgelei van voedsel (eksogene cholesterol). Die verwydering van cholesterol uit weefsels vind plaas deur die oksidasie daarvan tot galsure in die lewer, gevolg deur hul uitskeiding met ontlasting (ongeveer 0,5 g per dag) en deur die uitskeiding van onveranderde cholesterol (ook met ontlasting). In stilstaande toestand: (Cholesteroleinde + Cholesterolind) - (Cholesterolekskr + Galsureekskr) = 0 As hierdie balans versteur word, verander die konsentrasie van cholesterol in die weefsel en in die bloed. Verhoogde cholesterol in bloed - hipercholesterolemie. Dit verhoog die waarskynlikheid van aterosklerose en galsteensiekte. REGULASIE VAN LIPID-uitruilings Lipiedmetabolisme word deur die sentrale senuweestelsel gereguleer. Langdurig negatiewe emosionele spanning, kan 'n toename in die vrystelling van catecholamines in die bloedstroom merkbare gewigsverlies veroorsaak. effek glukagon op die lipolitiese stelsel is soortgelyk aan die werking van kategolamiene. adrenalien en norepinefrien verhoog die aktiwiteit van weefsellipase en die tempo van lipolise in vetweefsel, waardeur die inhoud van vetsure in bloedplasma toeneem. insulien het die teenoorgestelde effek van adrenalien en glukagon op lipolise en mobilisering van vetsure. Groeihormoon stimuleer lipolise, wat die sintese van asenilaat siklas veroorsaak. Hypofunksie van die hypofyse lei tot die afsetting van vet in die liggaam (vetsug in die pituïtêre liggaam). Tiroksien, geslagshormonebeïnvloed ook lipiedmetabolisme. Die verwydering van die geslagskliere by diere veroorsaak oortollige vetneerlegging. LIPID METABOLIESE VERSKIL Cholesterol is 'n steroïed wat spesifiek vir diereorganismes is. Die belangrikste plek waar dit in die menslike liggaam gevorm word, is die lewer, waar 50% cholesterol gesintetiseer word, 15-20% in die dunderm gevorm word, en die res in die vel, bynierskors en gonades gesintetiseer word. Die bronne van die vorming van die cholesterolfonds en die besteding daarvan word in Fig. 22.1 weergegee. Fig. 22.1. Die vorming en verspreiding van cholesterol in die liggaam. Die cholesterol van die menslike liggaam (totale hoeveelheid van ongeveer 140 g) kan voorwaardelik in drie poele verdeel word: 30 g), vinnig uitruilend, bestaan uit cholesterol van die dermwand, bloedplasma, lewer en ander parenchymale organe; hernuwing vind plaas in 30 dae (1 g / dag), 50 g), om cholesterol van ander organe en weefsels stadig uit te ruil, 60 g), 'n baie stadig uitgewisselde cholesterol van die rugmurg en brein, bindweefsel, word die opdateringskoers oor jare bereken. Die sintese van cholesterol vind plaas in die sitosol van selle. Dit is een van die langste metaboliese weë in die menslike liggaam. Dit gaan in drie fases voort: die eerste eindig met die vorming van mevalonzuur, die tweede met die vorming van squaleen (lineêre koolwaterstofstruktuur bestaande uit 30 koolstofatome). Gedurende die derde fase word squaleen omgeskakel in 'n lanosterolmolekule, en dan is daar 20 opeenvolgende reaksies wat lanosterol omskakel in cholesterol. In sommige weefsels verester die hidroksielgroep cholesterol om esters te vorm. Die reaksie word gekataliseer deur die intrasellulêre ensiem AHAT (acylCoA: cholesterol acyltransferase). Die esterifiseringsreaksie kom ook in die bloed voor in HDL, waar die ensiem LHAT (lesitien: cholesterol-asieltransferase) geleë is. Cholesterolesters is die vorm waarin dit deur bloed vervoer word of in selle neergesit word. In die bloed is ongeveer 75% cholesterol in die vorm van esters. Cholesterolsintese word gereguleer deur die aktiwiteit en hoeveelheid van die sleutelensiem van die proses te beïnvloed - 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA reductase). Dit word op twee maniere bereik: 1. Fosforylering / defosforylering van HMG-CoA-reduktase. Insulien stimuleer die defosforylering van HMG-CoA-reduktase en vertaal dit dan in 'n aktiewe toestand. Daarom neem die sintese van cholesterol in die absorpsietydperk toe. Gedurende hierdie periode neem die beskikbaarheid van die beginnersubstraat vir sintese, asetiel-CoA, ook toe. Glukagon het die teenoorgestelde effek: deur proteïenkinase A stimuleer dit die fosforylering van HMG-CoA-reduktase en word dit in 'n onaktiewe toestand. As gevolg hiervan word die sintese van cholesterol in die postabsorpsieperiode en tydens vas belemmer. 2. Inhibisie van die sintese van HMG-CoA reduktase.Cholesterol (eindproduk van die metaboliese weg) verminder die transkripsiesnelheid van die HMG-CoA-reduktase-geen, en belemmer dus die eie sintese, en galsure veroorsaak ook 'n soortgelyke effek. Die vervoer van bloedcholesterol word as deel van die middel uitgevoer. LP's bied eksogene cholesterol in weefsels, bepaal die vloei daarvan tussen organe en die uitskeiding van die liggaam. Eksogene cholesterol word as deel van die resterende ChM in die lewer afgegee. Daar vorm dit saam met gesintetiseerde endogene cholesterol 'n gemeenskaplike fonds. By hepatosiete word TAGs en cholesterol in VLDL verpak en in hierdie vorm in die bloed afgeskei. Onder die invloed van LP-lipase, wat TAG hydroliseer tot glycerol en vetsure in bloed, word VLDLP's eers omgeskakel na STLP's en dan na LDLP's wat tot 55% cholesterol en sy esters bevat. LDL is die vernaamste vervoervorm van cholesterol waarin dit aan weefsels oorgedra word (70% van die cholesterol en sy esters in die bloed vorm deel van LDL). LDL uit die bloed kom in die lewer (tot 75%) en ander weefsels wat LDL-reseptore op hul oppervlak het. As die hoeveelheid cholesterol wat in die sel binnekom, die behoefte daarvan oorskry, word die sintese van LDL-reseptore onderdruk, wat die vloei van cholesterol uit die bloed verminder. Inteendeel, dan word die reseptorsintese geaktiveer met 'n afname in die konsentrasie van vrye cholesterol in die sel. Hormone is betrokke by die regulering van LDL-reseptorsintese: insulien, triiodothyronine en geslagshormone verhoog die vorming van reseptore, en glukokortikoïede neem af. In die sogenaamde “cholesterol-terugvoer-vervoer”, d.w.s. die pad wat die terugkeer van cholesterol na die lewer verseker, speel HDL die hoofrol. Dit word in die lewer gesintetiseer in die vorm van onvolwasse voorgangers wat prakties nie cholesterol en TAG bevat nie. HDL-voorgangers in die bloed is versadig met cholesterol en ontvang dit van ander LP's en selmembrane. Die oordrag van cholesterol na HDL behels die LHAT-ensiem wat op hul oppervlak geleë is. Hierdie ensiem heg 'n vetsuurresidu van fosfatidielcholien (lesitien) aan cholesterol. As gevolg hiervan word 'n hidrofobiese molekule van cholesterolester gevorm wat binne HDL beweeg. Dus, nie dronk HDL, verryk met cholesterol nie, word dus HDL 3 - volwasse en groter deeltjies. HDL 3 ruil cholesterolesters vir TAG vervat in VLDL en STD met die deelname van 'n spesifieke proteïen wat cholesterolesters tussen lipoproteïne oordra. In hierdie geval, HDL 3 verander in HDL2, waarvan die grootte toeneem as gevolg van die ophoping van TAG. VLDL en STDL onder die invloed van LP-lipase word omgeskakel na LDL, wat hoofsaaklik cholesterol in die lewer lewer. 'N Klein hoeveelheid cholesterol word in die lewer van HDL2 en HDL afgegee. Sintese van galsure. In die lewer word 500–700 mg galsure per dag van cholesterol gesintetiseer. Hul vorming sluit reaksies in van die bekendstelling van hidroksielgroepe met die deelname van hidroksilase en die reaksie van gedeeltelike oksidasie van die syketting van cholesterol (Fig. 22.2): Fig. 22.2. Skema vir die vorming van galsure. Die eerste sintesiereaksie - die vorming van 7-a-hydroxycholesterol - is regulatories. Die aktiwiteit van die ensiem wat hierdie reaksie kataliseer, word belemmer deur die eindproduk van die baan, galsure. 'N Ander reguleringsmeganisme is fosforilering / defosforilering van die ensiem (die gefosforileerde vorm van 7-a-hidroksilase is aktief). Regulering is ook moontlik deur die hoeveelheid enzym te verander: cholesterol veroorsaak transkripsie van die 7-a-hidroksilase-geen, en galsure onderdruk. Skildklierhormone veroorsaak die sintese van 7-a-hidroksilase en estrogeenonderdrukking. So 'n effek van estrogeen op die sintese van galsure verklaar waarom galsteensiekte 3-4 keer meer gereeld by vroue voorkom as by mans. Choliese en chenodeoksigoliese sure wat gevorm word uit cholesterol, word “primêre galsure” genoem. Die grootste deel van hierdie sure word gekonjugeer - die toevoeging van glisien- of taurienmolekules tot die karboksielgroep galsuur. Konjugasie begin met die vorming van die aktiewe vorm van galsure - afgeleides van CoA, dan word taurien of glisien aangeheg, en as gevolg hiervan word 4 variante van konjugate gevorm: taurocholiese en taurohenodeoxycholiese, glikocholiese en glikohenodeoxicholiese sure. Dit is aansienlik sterker emulgeermiddels as die oorspronklike galsure. Konjugate met glisien word drie keer meer gevorm as met taurien, aangesien die hoeveelheid taurien in die liggaam beperk is. In die ingewande word 'n klein hoeveelheid konjugate van primêre galsure onder die werking van bakteriële ensieme omgesit in sekondêre galsure. Deoksicholiese suur, gevorm uit cholies, en litocholies, gevorm deur deoksicholies, is minder oplosbaar en word stadiger in die ingewande geabsorbeer. Ongeveer 95% van die galsure wat die ingewande binnekom, keer deur die porale aar na die lewer, en word dan weer in die gal geskei en weer gebruik in die emulsifisering van vette. Hierdie weg van galsure word enterohepatiese sirkulasie genoem. Met ontlasting word sekondêre galsure meestal verwyder. Gallsteensiekte (cholelithiasis) is 'n patologiese proses waarin klippe in die galblaas vorm, waarvan die basis cholesterol is. Die vrystelling van cholesterol in gal moet gepaard gaan met 'n proporsionele vrystelling van galsure en fosfolipiede wat hidrofobiese cholesterolmolekules in die micellêre toestand hou. Die redes wat lei tot 'n verandering in die verhouding van galsure en cholesterol in gal, is: voedsel ryk aan cholesterol, voeding met 'n hoë kalorie, stagnasie van gal in die galblaas, verswakte enterohepatiese sirkulasie, verswakte sintese van galsure, galblaasinfeksie. By die meeste pasiënte met cholelithiasis word die sintese van cholesterol verhoog, en die sintese van galsure daaruit vertraag, wat lei tot 'n onevenredigheid in die hoeveelheid cholesterol en galsure wat in die gal geskei word. As gevolg hiervan begin cholesterol in die galblaas neerslag en vorm dit 'n viskose neerslag wat geleidelik verhard. Soms word dit geïmpregneer met bilirubien, proteïene en kalsiumsoute. Stene kan slegs bestaan uit cholesterol (cholesterolstene) of 'n mengsel van cholesterol, bilirubien, proteïene en kalsium. Cholesterolstene is gewoonlik wit, en gemengde klippe is bruin in verskillende skakerings. In die eerste fase van die vorming van klip kan chenodeoxycholic suur as medisyne gebruik word. Sodra dit in die galblaas is, los dit die cholesterolstene geleidelik op, maar dit is 'n stadige proses wat etlike maande duur. Aterosklerose is 'n patologie wat gekenmerk word deur die voorkoms van aterogene gedenkplate op die binne-oppervlak van die vaskulêre wand. Een van die hoofredes vir die ontwikkeling van so 'n patologie is 'n skending van die balans tussen die inname van cholesterol uit voedsel, die sintese daarvan en die uitskeiding van die liggaam. Pasiënte met aterosklerose het verhoogde LDL- en VLDL-konsentrasies. Daar is 'n omgekeerde verband tussen HDL-konsentrasie en die waarskynlikheid dat aterosklerose ontwikkel. Dit stem ooreen met die konsep van die funksionering van LDL as draers van cholesterol in weefsels en HDL in weefsels. Die basiese metaboliese "voorvereiste" vir die ontwikkeling van aterosklerose is hipercholesterolemie. (hoë cholesterol in die bloed). Hipercholesterolemie ontwikkel: 1. as gevolg van 'n oormatige inname van cholesterol, koolhidrate en vette, 2. 'n genetiese geneigdheid wat bestaan uit oorerflike afwykings in die struktuur van LDL-reseptore of apoB-100, sowel as in 'n verhoogde sintese of afskeiding van apoB-100 (in die geval van familiële gekombineerde hiperlipidemie, waarin bloedkonsentrasies en cholesterol en TAG verhoog word). 'N Belangrike rol in die meganismes vir die ontwikkeling van aterosklerose word gespeel deur die modifisering van die geneesmiddel. Veranderings in die normale struktuur van lipiede en proteïene in LDL maak dit vreemd vir die liggaam en daarom meer toeganklik vir opname deur fagosiete. Medikasie-verandering kan deur verskeie meganismes plaasvind: 1. glikosilering van proteïene wat plaasvind wanneer die konsentrasie van glukose in die bloed toeneem, 2. peroksiedmodifikasie, wat lei tot veranderinge in lipiede in lipoproteïene en die struktuur van apoB-100, 3. die vorming van outo-immuunkomplekse van die LP-teenliggaampie (veranderde medisyne kan die vorming van auto-antiliggame veroorsaak). Gemodifiseerde LDL word geabsorbeer deur makrofage. Hierdie proses word nie gereguleer deur die hoeveelheid cholesterol wat geabsorbeer word nie, soos in die geval van seleksie deur spesifieke reseptore, en daarom word makrofage oorlaai met cholesterol en word dit 'skuimagtige selle' wat in die subendoteliale ruimte binnedring. Dit lei tot die vorming van lipied kolle of stroke in die wand van bloedvate. Op hierdie stadium kan die vaskulêre endoteel sy struktuur behou. Met 'n toename in die aantal skuimselle, vind endotheelskade plaas. Skade dra by tot die aktivering van plaatjies. As gevolg hiervan skei hulle tromboksaan af, wat plaatjie-aggregasie stimuleer, en begin ook plaatjie-groeifaktor produseer, wat die verspreiding van gladdespierselle stimuleer. Laasgenoemde migreer van die mediale na die binneste laag van die arteriële wand, en dra dus by tot die groei van die gedenkplaat. Verder spruit die gedenkplaat met veselagtige weefsel, die selle onder die veselagtige membraan is nekroties, en cholesterol word in die intersellulêre ruimte neergesit. In die laaste stadiums van ontwikkeling word die gedenkplaat met kalsiumsoute geïmpregneer en word dit baie dig. In die area van die gedenkplaat vorm daar gereeld bloedklonte wat die lumen van die vaart versper, wat lei tot akute versteuring van die bloedsomloop in die ooreenstemmende weefselplek en die ontwikkeling van 'n hartaanval. Die regulering van die sintese van cholesterol - die sleutelensiem daarvan (HMG-CoA-reduktase) word op verskillende maniere uitgevoer. Fosforylering / defosforylering van HMG-reduktase. Met die toename in die insulien / glukagon-verhouding fosforylateer hierdie ensiem en beweeg dit in 'n aktiewe toestand. Die werking van insulien word deur 2 ensieme uitgevoer. HMG-CoA reduktase kinase fosfatase, wat die kinase omskep in 'n onaktiewe gedefosforileerde toestand: Fosfotase HMG-CoA reduktase deur dit in 'n gedefosforyleerde aktiewe toestand om te skakel. Die resultaat van hierdie reaksies is die vorming van 'n gedefosforyleerde aktiewe vorm van HMG-CoA-reduktase. Gevolglik neem cholesterol toe gedurende die absorpsietydperk. Gedurende hierdie periode neem die beskikbaarheid van die aanvanklike substraat vir die sintese van cholesterol - asetiel-CoA toe (as gevolg van die eet van voedsel wat koolhidrate en vette bevat, aangesien CoA-asetiel gevorm word tydens die afbreek van glukose en vetsure). In die postabsorberende toestand stimuleer glukagon deur proteingenase A die fosforylering van HMG - CoA - reduktase, wat dit in 'n onaktiewe toestand maak. Hierdie werking word versterk deurdat glukagon terselfdertyd die fosforilering en die inaktivering van die fosfotase van HMG-CoA-reduktase stimuleer, en sodoende die HMG-CoA-reduktase in 'n gefosforyleerde onaktiewe toestand gehou word. As gevolg hiervan, word die sintese van cholesterol in die postabsorpsieperiode en tydens vas geïnhibeer deur endogene sintese. As die cholesterolinhoud in voedsel op 2% gebring word, neem die sintese van endogene cholesterol skerp af. Maar die volledige staking van cholesterol-sintese kom nie voor nie. Die mate van inhibisie van cholesterolbiosintese onder invloed van cholesterol wat van voedsel afkomstig is, wissel van persoon tot persoon. Dit dui op die individualiteit van die prosesse van cholesterolvorming. Deur die intensiteit van cholesterolsintese te verminder, is dit moontlik om die konsentrasie in die bloed te verminder. As die balans tussen die inname van cholesterol met voedsel en die sintese daarvan in die liggaam aan die een kant en die uitskeiding van galsure en cholesterol aan die ander kant verbreek word, verander die konsentrasie van cholesterol in die weefsel en bloed. Die ernstigste gevolge hou verband met 'n toename in bloedkolesterolkonsentrasie (hipercholesterolemie), terwyl die waarskynlikheid om aterosklerose en cholelithiasis te ontwikkel, verhoog. Gesinshipercholesterolemie (HCS) - hierdie vorm kom baie meer voor - ongeveer 1 pasiënt per 200 mense. 'N Geërfde defek in HCS is 'n skending van die opname van LDL deur selle, en dus 'n afname in die tempo van LDL-katabolisme. As gevolg hiervan styg die konsentrasie van LDL in die bloed, sowel as cholesterol, aangesien daar baie daarvan in LDL is. Daarom is die afsetting van cholesterol in weefsels, veral in die vel (xanthomas), in die wande van arteries kenmerkend met HCS. Inhibering van die sintese van HMG-CoA-reduktase Die finale produk van die metaboliese metaboliese weg. Dit verminder die tempo van transkripsie van die HMG-CoA-reduktase-geen, en belemmer dus sy eie sintese. Die lewer sintetiseer galsure aktief van cholesterol, en die remming van die galsure is dus die aktiwiteit van die HMG-CoA-reduktase-geen. Aangesien HMG-CoA-reduktase na sintese van ongeveer 3 bestaan, is die inhibering van die sintese van hierdie cholesterolensiem 'n effektiewe regulering. Kyk na die video: Celstofwisseling - regulering van de enzymactiviteit - VWO (November 2024).
|